

Which Of The Following Is Not A Characteristic Of An Animal Plasma Membrane
Like all other cellular membranes, the plasma membrane consists of both lipids and proteins. The primal structure of the membrane is the phospholipid bilayer, which forms a stable barrier between 2 aqueous compartments. In the case of the plasma membrane, these compartments are the inside and the outside of the prison cell. Proteins embedded within the phospholipid bilayer bear out the specific functions of the plasma membrane, including selective transport of molecules and cell-cell recognition.
The Phospholipid Bilayer
The plasma membrane is the most thoroughly studied of all cell membranes, and it is largely through investigations of the plasma membrane that our current concepts of membrane structure accept evolved. The plasma membranes of mammalian red claret cells (erythrocytes) have been particularly useful as a model for studies of membrane structure. Mammalian ruddy blood cells practice not contain nuclei or internal membranes, and then they represent a source from which pure plasma membranes can be easily isolated for biochemical analysis. Indeed, studies of the red blood cell plasma membrane provided the commencement evidence that biological membranes consist of lipid bilayers. In 1925, two Dutch scientists (E. Gorter and R. Grendel) extracted the membrane lipids from a known number of cherry blood cells, corresponding to a known surface expanse of plasma membrane. They and so determined the surface area occupied past a monolayer of the extracted lipid spread out at an air-water interface. The area of the lipid monolayer turned out to be twice that occupied by the erythrocyte plasma membranes, leading to the conclusion that the membranes consisted of lipid bilayers rather than monolayers.
The bilayer structure of the erythrocyte plasma membrane is clearly evident in high-magnification electron micrographs (Effigy 12.1). The plasma membrane appears as ii dense lines separated by an intervening space—a morphology frequently referred to as a "railroad track" advent. This image results from the bounden of the electron-dense heavy metals used as stains in transmission electron microscopy (see Chapter 1) to the polar head groups of the phospholipids, which therefore appear as night lines. These dense lines are separated past the lightly stained interior portion of the membrane, which contains the hydrophobic fat acrid chains.

Effigy 12.1
Bilayer structure of the plasma membrane. Electron micrograph of a homo reddish claret cell. Note the railroad track appearance of the plasma membrane. (Courtesy of J. David Robertson, Knuckles University Medical Centre.)
As discussed in Chapter two, the plasma membranes of creature cells incorporate four major phospholipids (phosphatidylcholine, phosphatidylethanolamine, phosphatidylserine, and sphingomyelin), which together account for more one-half of the lipid in well-nigh membranes. These phospholipids are asymmetrically distributed betwixt the two halves of the membrane bilayer (Figure 12.2). The outer leaflet of the plasma membrane consists mainly of phosphatidylcholine and sphingomyelin, whereas phosphatidylethanolamine and phosphatidylserine are the predominant phospholipids of the inner leaflet. A 5th phospholipid, phosphatidylinositol, is also localized to the inner one-half of the plasma membrane. Although phosphatidylinositol is a quantitatively minor membrane component, information technology plays an important function in cell signaling, every bit discussed in the adjacent chapter. The head groups of both phosphatidylserine and phosphatidylinositol are negatively charged, so their predominance in the inner leaflet results in a net negative charge on the cytosolic face up of the plasma membrane.
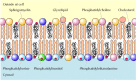
Effigy 12.ii
Lipid components of the plasma membrane. The outer leaflet consists predominantly of phosphatidylcholine, sphingomyelin, and glycolipids, whereas the inner leaflet contains phosphatidylethanolamine, phosphatidylserine, and phosphatidylinositol. Cholesterol (more...)
In addition to the phospholipids, the plasma membranes of brute cells contain glycolipids and cholesterol. The glycolipids are constitute exclusively in the outer leaflet of the plasma membrane, with their carbohydrate portions exposed on the cell surface. They are relatively minor membrane components, constituting only about 2% of the lipids of near plasma membranes. Cholesterol, on the other hand, is a major membrane elective of brute cells, being present in nigh the same molar amounts as the phospholipids.
Ii general features of phospholipid bilayers are critical to membrane function. Outset, the construction of phospholipids is responsible for the bones function of membranes every bit barriers between two aqueous compartments. Because the interior of the phospholipid bilayer is occupied past hydrophobic fatty acrid chains, the membrane is impermeable to water-soluble molecules, including ions and most biological molecules. Second, bilayers of the naturally occurring phospholipids are viscous fluids, non solids. The fatty acids of most natural phospholipids take one or more double bonds, which innovate kinks into the hydrocarbon chains and make them difficult to pack together. The long hydrocarbon chains of the fat acids therefore move freely in the interior of the membrane, and so the membrane itself is soft and flexible. In addition, both phospholipids and proteins are free to diffuse laterally inside the membrane—a property that is disquisitional for many membrane functions.
Because of its rigid ring construction, cholesterol plays a distinct role in membrane structure. Cholesterol will not form a membrane by itself, just inserts into a bilayer of phospholipids with its polar hydroxyl group close to the phospholipid head groups (encounter Figure 12.2). Depending on the temperature, cholesterol has distinct effects on membrane fluidity. At high temperatures, cholesterol interferes with the motility of the phospholipid fat acrid bondage, making the outer part of the membrane less fluid and reducing its permeability to small molecules. At depression temperatures, however, cholesterol has the opposite effect: By interfering with interactions betwixt fatty acid bondage, cholesterol prevents membranes from freezing and maintains membrane fluidity. Although cholesterol is not nowadays in leaner, information technology is an essential component of animal cell plasma membranes. Plant cells also lack cholesterol, but they contain related compounds (sterols) that fulfill a similar part.
Recent studies propose that not all lipids diffuse freely in the plasma membrane. Instead, detached membrane domains appear to be enriched in cholesterol and the sphingolipids (sphingomyelin and glycolipids). These clusters of sphingolipids and cholesterol are thought to class "rafts" that motility laterally within the plasma membrane and may associate with specific membrane proteins. Although the functions of lipid rafts remain to be understood, they may play of import roles in processes such as cell signaling and the uptake of extracellular molecules by endocytosis.
Membrane Proteins
While lipids are the fundamental structural elements of membranes, proteins are responsible for carrying out specific membrane functions. Most plasma membranes consist of approximately l% lipid and 50% poly peptide by weight, with the carbohydrate portions of glycolipids and glycoproteins constituting five to 10% of the membrane mass. Since proteins are much larger than lipids, this percentage corresponds to about 1 protein molecule per every 50 to 100 molecules of lipid. In 1972, Jonathan Vocalist and Garth Nicolson proposed the fluid mosaic model of membrane structure, which is at present by and large accepted as the bones paradigm for the organisation of all biological membranes. In this model, membranes are viewed as two-dimensional fluids in which proteins are inserted into lipid bilayers (Figure 12.3).

Figure 12.3
Fluid mosaic model of the plasma membrane. Integral membrane proteins are inserted into the lipid bilayer, whereas peripheral proteins are leap to the membrane indirectly by poly peptide-protein interactions. Most integral membrane proteins are transmembrane (more than...)
Vocalizer and Nicolson distinguished two classes of membrane-associated proteins, which they called peripheral and integral membrane proteins. Peripheral membrane proteins were operationally defined as proteins that dissociate from the membrane following treatments with polar reagents, such as solutions of extreme pH or high salt concentration, that practise not disrupt the phospholipid bilayer. Once dissociated from the membrane, peripheral membrane proteins are soluble in aqueous buffers. These proteins are not inserted into the hydrophobic interior of the lipid bilayer. Instead, they are indirectly associated with membranes through protein-protein interactions. These interactions frequently involve ionic bonds, which are disrupted past extreme pH or loftier salt.
In contrast to the peripheral membrane proteins, integral membrane proteins can exist released only by treatments that disrupt the phospholipid bilayer. Portions of these integral membrane proteins are inserted into the lipid bilayer, then they can exist dissociated only past reagents that disrupt hydrophobic interactions. The most commonly used reagents for solubilization of integral membrane proteins are detergents, which are small amphipathic molecules containing both hydrophobic and hydrophilic groups (Effigy 12.4). The hydrophobic portions of detergents displace the membrane lipids and bind to the hydrophobic portions of integral membrane proteins. Because the other terminate of the detergent molecule is hydrophilic, the detergent-protein complexes are soluble in aqueous solutions.

Figure 12.4
Solubilization of integral membrane proteins by detergents. Detergents (e.g., octyl glucoside) are amphipathic molecules containing hydrophilic head groups and hydrophobic tails. The hydrophobic tails bind to the hydrophobic regions of integral membrane (more...)
Many integral proteins are transmembrane proteins, which bridge the lipid bilayer with portions exposed on both sides of the membrane. These proteins can be visualized in electron micrographs of plasma membranes prepared by the freeze-fracture technique (see Figure 1.35). In these specimens, the membrane is split and separates into its two leaflets. Transmembrane proteins are then credible as particles on the internal faces of the membrane (Figure 12.5).

Figure 12.v
Freeze-fracture electron micrograph of human being cerise blood jail cell membranes. The particles in the membrane are transmembrane proteins. (Harold H. Edwards/Visuals Unlimited.)
The membrane-spanning portions of transmembrane proteins are usually α helices of 20 to 25 hydrophobic amino acids that are inserted into the membrane of the endoplasmic reticulum during synthesis of the polypeptide chain (see Figures 9.11, 9.12, and ix.13). These proteins are then transported in membrane vesicles from the endoplasmic reticulum to the Golgi apparatus, and from there to the plasma membrane. Carbohydrate groups are added to the polypeptide chains in both the endoplasmic reticulum and Golgi apparatus, so well-nigh transmembrane proteins of the plasma membrane are glycoproteins with their oligosaccharides exposed on the surface of the prison cell.
Studies of red claret cells have provided good examples of both peripheral and integral proteins associated with the plasma membrane. The membranes of human erythrocytes contain almost a dozen major proteins, which were originally identified past gel electrophoresis of membrane preparations. Almost of these are peripheral membrane proteins that accept been identified as components of the cortical cytoskeleton, which underlies the plasma membrane and determines cell shape (run into Chapter eleven). For case, the most abundant peripheral membrane poly peptide of ruby-red blood cells is spectrin, which is the major cytoskeletal protein of erythrocytes. Other peripheral membrane proteins of carmine claret cells include actin, ankyrin, and band 4.1. Ankyrin serves as the principal link betwixt the plasma membrane and the cytoskeleton by binding to both spectrin and the integral membrane poly peptide band 3 (meet Figure 11.11). An boosted link between the membrane and the cytoskeleton is provided by band four.one, which binds to the junctions of spectrin and actin, as well as to glycophorin (the other major integral membrane protein of erythrocytes).
The ii major integral membrane proteins of cerise claret cells, glycophorin and band 3, provide well-studied examples of transmembrane protein structure (Figure 12.6). Glycophorin is a modest glycoprotein of 131 amino acids, with a molecular weight of about xxx,000, one-half of which is protein and half carbohydrate. Glycophorin crosses the membrane with a unmarried membrane-spanning α helix of 23 amino acids, with its glycosylated amino-final portion exposed on the cell surface. Although glycophorin was one of the first transmembrane proteins to exist characterized, its precise office remains unknown. In contrast, the function of the other major transmembrane protein of red blood cells is well understood. This protein, originally known as band 3, is the anion transporter responsible for the passage of bicarbonate (HCO3 -) and chloride (Cl-) ions beyond the red claret jail cell membrane. The band 3 polypeptide chain is 929 amino acids and is thought to take 14 membrane-spanning α-helical regions. Within the membrane, dimers of band three form globular structures containing internal channels through which ions are able to travel beyond the lipid bilayer.
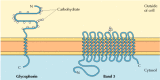
Figure 12.half-dozen
Integral membrane proteins of ruddy blood cells. Glycophorin (131 amino acids) contains a unmarried transmembrane α helix. It is heavily glyocosylated, with oligosaccharides fastened to sixteen sites on the extracellular portion of the polypeptide chain. (more...)
Because of their amphipathic character, transmembrane proteins have proved difficult to crystallize, as required for iii-dimensional structural analysis by 10-ray diffraction. The first transmembrane poly peptide to be analyzed by X-ray crystallography was the photosynthetic reaction center of the bacterium Rhodopseudomonas viridis, whose structure was reported in 1985 (Figure 12.7). The reaction center contains three transmembrane proteins, designated L, M, and H (light, medium, and heavy) according to their apparent sizes indicated by gel electrophoresis. The L and M subunits each have 5 membrane-spanning α helices. The H subunit has just a single transmembrane α helix, with the majority of the polypeptide chain on the cytosolic side of the membrane. The quaternary subunit of the reaction heart is a cytochrome, which is a peripheral membrane poly peptide leap to the complex by protein-protein interactions.

Figure 12.7
A bacterial photosynthetic reaction eye. The reaction center consists of three transmembrane proteins, designated L (red), M (yellow), and H (light-green). The L and M subunits each have five transmembrane α helices, whereas the H subunit has simply (more...)
Although nigh transmembrane proteins span the membrane by α-helical regions, this is not always the case. A well-characterized exception is provided past the porins—a class of proteins that form channels in the outer membranes of some bacteria. Many leaner, including E. coli, have a dual membrane system in which the plasma membrane (or inner membrane) is surrounded by the jail cell wall and a distinct outer membrane (Figure 12.viii). In contrast to the plasma membrane, the outer membrane is highly permeable to ions and small-scale polar molecules (in the instance of E. coli, with molecular weights upward to 600). This permeability results from the porins, which form open aqueous channels through the lipid bilayer. As discussed in Affiliate x, proteins related to the bacterial porins are also found in the outer membranes of mitochondria and chloroplasts.
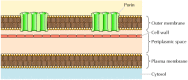
Figure 12.8
Bacterial outer membranes. The plasma membrane of some leaner is surrounded past a cell wall and a distinct outer membrane. The outer membrane contains porins, which form open aqueous channels allowing the gratis passage of ions and small molecules.
Structural analysis has indicated that the porins practise not incorporate hydrophobic α-helical regions. Instead, they cantankerous the membrane as β barrels, in which 16 β sheets fold up into a barrel-like structure enclosing an aqueous pore (Figure 12.nine). The side chains of polar amino acids line the pore, whereas side chains of hydrophobic amino acids interact with the interior of the membrane. The porin monomers acquaintance to form stable trimers, each of which contains three open channels through which polar molecules can diffuse beyond the membrane.

Figure 12.9
Structure of a porin monomer. Each monomer is a β barrel consisting of 16 antiparallel β strands (arrows). The top end of the molecule faces the external medium. (From H. Nikaido, 1994. J. Biol. Chem. 269: 3905.)
In contrast to transmembrane proteins, a multifariousness of proteins (many of which acquit as integral membrane proteins) are anchored in the plasma membrane past covalently attached lipids or glycolipids (Figure 12.x). Members of one form of these proteins are inserted into the outer leaflet of the plasma membrane past glycosylphosphatidylinositol (GPI) anchors. GPI anchors are added to sure proteins that have been transferred into the endoplasmic reticulum and are anchored in the membrane by a C-terminal transmembrane region (run across Figure ix.16). The transmembrane region is cleaved as the GPI ballast is added, and so these proteins remain attached to the membrane only by the glycolipid. Since the polypeptide chains of GPI-anchored proteins are transferred into the endoplasmic reticulum, they are glycosylated and exposed on the surface of the jail cell following transport to the plasma membrane.
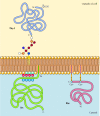
Figure 12.10
Examples of proteins anchored in the plasma membrane past lipids and glycolipids. Some proteins (e.g., the lymphocyte protein Thy-1) are anchored in the outer leaflet of the plasma membrane by GPI anchors added to their C terminus in the endoplasmic reticulum. (more...)
Other proteins are anchored in the inner leaflet of the plasma membrane by covalently fastened lipids. Rather than being candy through the secretory pathway, these proteins are synthesized on free cytosolic ribosomes and and so modified by the improver of lipids. These modifications include the addition of myristic acrid (a xiv-carbon fatty acid) to the amino terminus of the polypeptide chain, the add-on of palmitic acid (16 carbons) to the side chains of cysteine residues, and the addition of prenyl groups (15 or 20 carbons) to the side chains of carboxy-last cysteine residues (see Figures vii.29, vii.thirty, and vii.31). In some cases, these proteins (many of which conduct as peripheral membrane proteins) are targeted to the plasma membrane by positively charged regions of the polypeptide chain as well equally past the attached lipids. These positively charged protein domains may interact with the negatively charged head groups of phosphatidylserine on the cytosolic confront of the plasma membrane. It is noteworthy that many of the proteins anchored in the inner leaflet of the plasma membrane (including the Src and Ras proteins illustrated in Figure 12.10) play important roles in the transmission of signals from prison cell surface receptors to intracellular targets, equally discussed in the side by side chapter.
Mobility of Membrane Proteins
Membrane proteins and phospholipids are unable to move back and along between the inner and outer leaflets of the membrane at an observable rate. Nevertheless, because they are inserted into a fluid lipid bilayer, both proteins and lipids are able to lengthened laterally through the membrane. This lateral motility was first shown directly in an experiment reported by Larry Frye and Michael Edidin in 1970, which provided back up for the fluid mosaic model. Frye and Edidin fused human and mouse cells in civilisation to produce human-mouse cell hybrids (Figure 12.11). They then analyzed the distribution of proteins in the membranes of these hybrid cells using antibodies that specifically recognize proteins of human and mouse origin. These antibodies were labeled with different fluorescent dyes, and then the homo and mouse proteins could exist distinguished by fluorescence microscopy. Immediately after fusion, human and mouse proteins were localized to different halves of the hybrid cells. Yet, subsequently a brief menstruum of incubation at 37°C, the human and mouse proteins were completely intermixed over the cell surface, indicating that they moved freely through the plasma membrane.

Effigy 12.11
Mobility of membrane proteins. Human and mouse cells were fused to produce hybrid cells. The distribution of cell surface proteins was then analyzed using anti-human and anti-mouse antibodies labeled with different fluorescent dyes (red and green, respectively). (more...)
However, not all proteins are able to diffuse freely through the membrane. In some cases, the mobility of membrane proteins is restricted by their clan with the cytoskeleton. For case, a fraction of band 3 in the red claret cell membrane is immobilized as a issue of its association with ankyrin and spectrin. In other cases, the mobility of membrane proteins may be restricted by their associations with other membrane proteins, with proteins on the surface of adjacent cells, or with the extracellular matrix.
In contrast to blood cells, epithelial cells are polarized when they are organized into tissues, with dissimilar parts of the cell responsible for performing distinct functions. Consequently, the plasma membranes of many epithelial cells are divided into distinct apical and basolateral domains that differ in function and protein composition (Figure 12.12). For instance, epithelial cells of the pocket-sized intestine office to absorb nutrients from the digestive tract. The upmost surface of these cells, which faces the intestinal lumen, is therefore covered by microvilli and specialized for nutrient assimilation. The basolateral surface, which faces underlying connective tissue and the claret supply, is specialized to mediate the transfer of absorbed nutrients into the circulation. In club to maintain these distinct functions, the mobility of plasma membrane proteins must be restricted to the appropriate domains of the cell surface. At to the lowest degree part of the machinery past which this occurs involves the germination of tight junctions (which are discussed later in this affiliate) between side by side cells of the epithelium. These junctions not only seal the space betwixt cells but also serve as barriers to the move of membrane lipids and proteins. As a result, proteins are able to diffuse within either the apical or basolateral domains of the plasma membrane simply are not able to cross from one domain to the other.
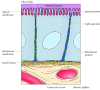
Figure 12.12
A polarized intestinal epithelial cell. The apical surface of the prison cell contains microvilli and is specialized for absorption of nutrients from the intestinal lumen. The basolateral surface is specialized for the transfer of absorbed nutrients to the underlying (more...)
The Glycocalyx
As already discussed, the extracellular portions of plasma membrane proteins are more often than not glycosylated. Besides, the sugar portions of glycolipids are exposed on the outer confront of the plasma membrane. Consequently, the surface of the jail cell is covered by a carbohydrate coat, known every bit the glycocalyx, formed by the oligosaccharides of glycolipids and transmembrane glycoproteins (Figure 12.thirteen).

Figure 12.thirteen
The glycocalyx. An electron micrograph of intestinal epithelium illustrating the glycocalyx (arrows). (Don Fawcett/ Visuals Unlimited.)
Part of the role of the glycocalyx is to protect the prison cell surface. In addition, the oligosaccharides of the glycocalyx serve as markers for a variety of jail cell-cell interactions. A well-studied case of these interactions is the adhesion of white claret cells (leukocytes) to the endothelial cells that line blood vessels—a process that allows the leukocytes to leave the circulatory system and mediate the inflammatory response in injured tissues. The initial footstep in adhesion betwixt leukocytes and endothelial cells is mediated past a family of transmembrane proteins called selectins, which recognize specific carbohydrates on the cell surface (Figure 12.14). 2 members of the selectin family (Eastward-selectin and P-selectin), expressed by endothelial cells and platelets, bind to specific oligosaccharides expressed on the surface of leukocytes. A different selectin (L-selectin) is expressed by leukocytes and recognizes an oligosaccharide on the surface of endothelial cells. The oligosaccharides exposed on the prison cell surface thus provide a set of markers that assistance place the distinct cell types of multicellular organisms.
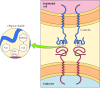
Figure 12.xiv
Binding of selectins to oligosaccharides. East-selectin is a transmembrane poly peptide expressed past endothelial cells that binds to an oligosaccharide expressed on the surface of leukocytes. The oligosaccharide recognized by Eastward-selectin contains N-acetylglucosamine (more...)
Source: https://www.ncbi.nlm.nih.gov/books/NBK9898/
Posted by: winklerwhadminvabot.blogspot.com
0 Response to "Which Of The Following Is Not A Characteristic Of An Animal Plasma Membrane"
Post a Comment